andrey khorst
andrey khorst
580 California St., Suite 400
San Francisco, CA, 94104
The book "Biochemical Pathways" aims to provide concise and comprehensive information on metabolic pathways, enzymatic reactions, and their regulation, without the extensive historical context typically found in textbooks. It focuses on a variety of biological systems, including bacteria, plants, and animals, presenting knowledge through clear illustrations and tables to facilitate quick understanding. The work is designed for readers looking for a systematic overview of biochemical interrelationships while omitting detailed experimental methods and the complete range of literature references.













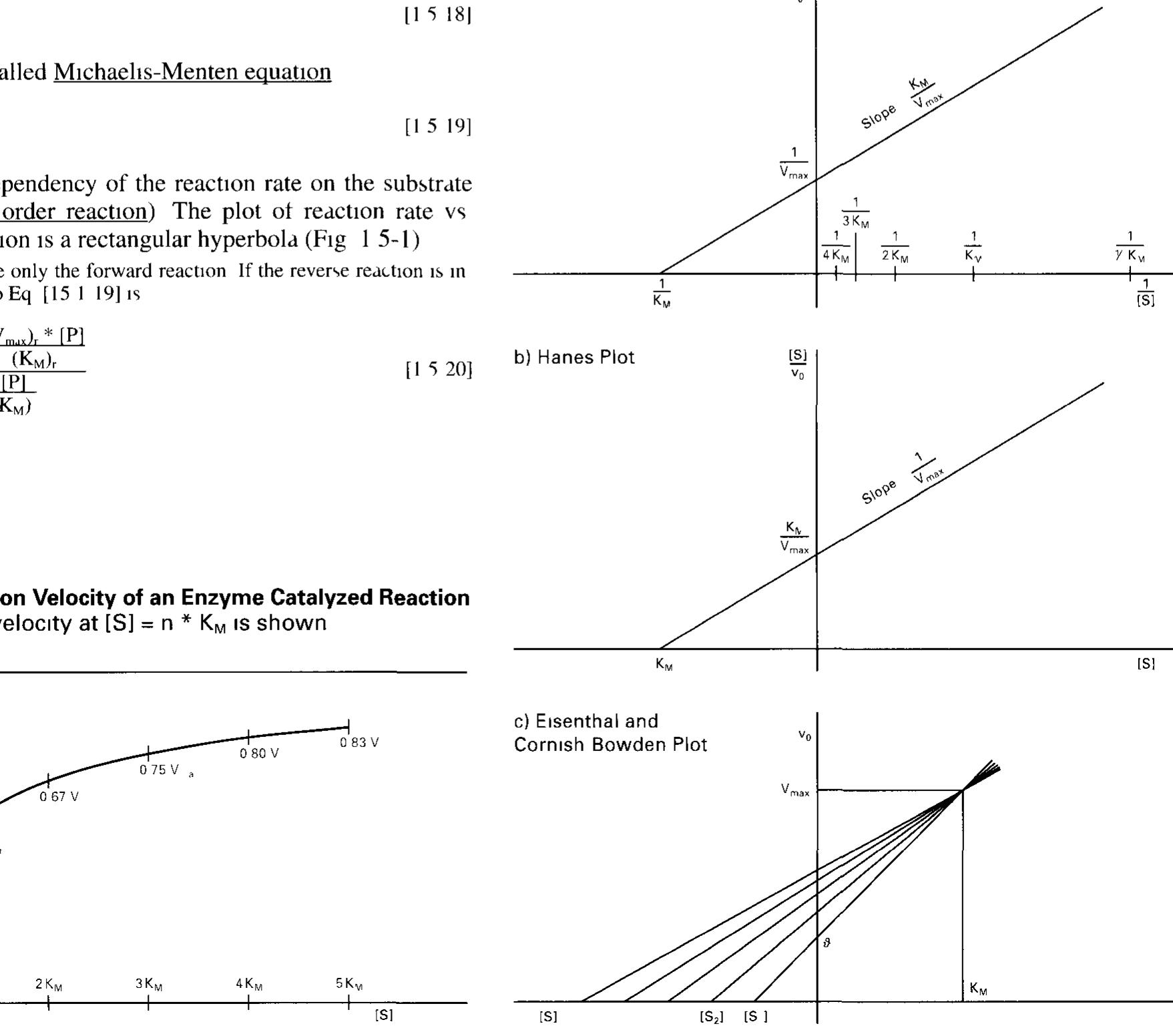

![The energy required tor its formation 1s called activation energy AG* which can be calculated from this equilibrium by applying Eq [1 5 4] as Arrow = shift of the plot when the concentration of the other substrate is raised “igure 1 5-5 Lineweaver-Burk Plots of Two-Substrate Reactions](https://figures.academia-assets.com/39117587/figure_009.jpg)

















![2.5.2 Regulation of the Activity of Enzymes (For a Mathematical Treatment see 1.5.4) Dannlatian danandinag am onhocteaota Camneantratinn«s TA tha mance? SECACHICIIE SOO 1.0. F) Regulation depending on substrate concentration: In the most simple case, the dependency of the reaction rate on the substrate concentration according to the Michaelis-Menten equation (Eq. [1.5-19], Fig. 1.5-1) 1s a means of controlling the substrate through- put. Zymogen activation: Some enzymes are synthesized as mactive precursors (zymogens or proenzymes) which have to be processed into their active form by limited proteolysis. Examples are diges- tive enzymes such as chymotrypsinogen or trypsinogen (Fig. 2.5-1) They are synthesized in mammalian pancreas as zymogens. After hormone-controlled release into the small intestine they are irre- versibly processed by trypsin or enteropeptidase, respectively, to become active proteases (Fig. 17.1-11).](https://figures.academia-assets.com/39117587/figure_022.jpg)
































































![Using [Mb ] for the total myoglobin concentration [Mb] + |Mb O ] a derivation analogous to the Michaelis Menten equation [Eq 15 19] but without further turnover to products yields](https://figures.academia-assets.com/39117587/figure_087.jpg)






![Figure 6.1-3. Schematic Drawing of the Animal Fatty Acid Synthase Dimer Reaction sequence (Fig. 6.1-4): Specific acy] transferases (one in animals, two in yeast) transfer an acetyl residue from acetyl-CoA to a cysteine-SH of the 3-oxoacyl synthase component (‘peripheral SH group’) @ and a malonyl residue to the ‘central’ SH group of the ACP component of the multienzyme system. Condensation with an acetoacetyl chain (bound to the central SH group) occurs with simultaneous decarboxylation, thereby energetically favoring chain elongation @). This step is irreversible. Then the resulting C, residue is reduced by NADPH at the oxoacyl reductase component ©, dehydrated to a 2,3-desaturated intermediate @ and reduced again by the enoyl reductase ® (also using NADPH as a reduc- tant; NADH in E.coli). The yeast enzyme contains flavin. Finally, the butyryl chain is transferred to the ‘peripheral’ SH group ©. Condensation with another malonyl! residue initiates another round of chain elongation.](https://figures.academia-assets.com/39117587/figure_091.jpg)






















![Farnesylation takes place if X = serine methionine or glutamine Geranylgerany] 1s attached 1f X = leucine or phenylalanine By Pane EDN ay ar HARES pp eh SMMC DON OSL t ASemmnO RODS BR ce a ee Re is pee Many proteins are attached to cellular membranes by isoprenoid anchors (via a thioether bond to mostly farnesyl, sometimes ge- ranylgeranyl chains), e g Ras (175), G, proteins (17 4) and heme A (521) This prenylation determines the location and the func tron of the proteins The enzymatic reaction 1s shown 1n Fig 7 3-1 IS AUACTIEM TE ZN = IRGC OF POUCH Y taal Proteins of the Rab group which contain -C-C- or C X C-~ sequences ar geranylgeranylated similarly The enzyme 1s assisted by the Rep protein](https://figures.academia-assets.com/39117587/figure_110.jpg)

























































![ee nnn ee EN I NEI SAE ROE NESE! Only a few regulatory events during the elongation step are known so far The overall RNA chain elongation rate, however, 1s propor tional to the growth rate, ensuring that no excess RNA 1s produced when the translation system operates slowly under conditions of liamited nutritional supply Stringent control: Starvation for an amino acid in bacteria causes a drastic decrease im transcription levels, caused by a signal from the translation step In this stringent contro] mechanism, translat ing ribosomes whose A sites are occupied by non-aminoacylated tRNAs bind the stringent factor enzyme, which then synthesizes the unusual nucleotide guanosine tetraphosphate (PP 5’ G 2’-PP) This compound, 1n turn, reduces transcription of rRNA and tRNA genes (and others) 10- to 20-fold by decreasing the affinity of Pol to the respective promoters via an unresolved mechanism On the other hand, amino acid biosynthesis 1s stimulated](https://figures.academia-assets.com/39117587/figure_162.jpg)


























![Figure 11 3-4 Splicing Mechanism for mRNA Besides this mechanism (group III introns) other splicing mechanisms exist They proceed with an external guanosine nucleotide instead of the branching point A (group I rRNA) or with a hgation procedure after endonuclease split ting of the pre mRNA (group [IV tRNA) Splicing of group II introns (e g muito chondria and chloroplast mRNA) resembles the group If] mechanism but with out participation of snRNPs Frequently the group I or I procedures are per tormed by RNA activity only (self splicing e g rRNA in Tetrahymena) Exon ot ditterent RNA strands can also be combined by splicing mechanisms (trans splicing e g MRNA 1n Trypanosoma)](https://figures.academia-assets.com/39117587/figure_180.jpg)





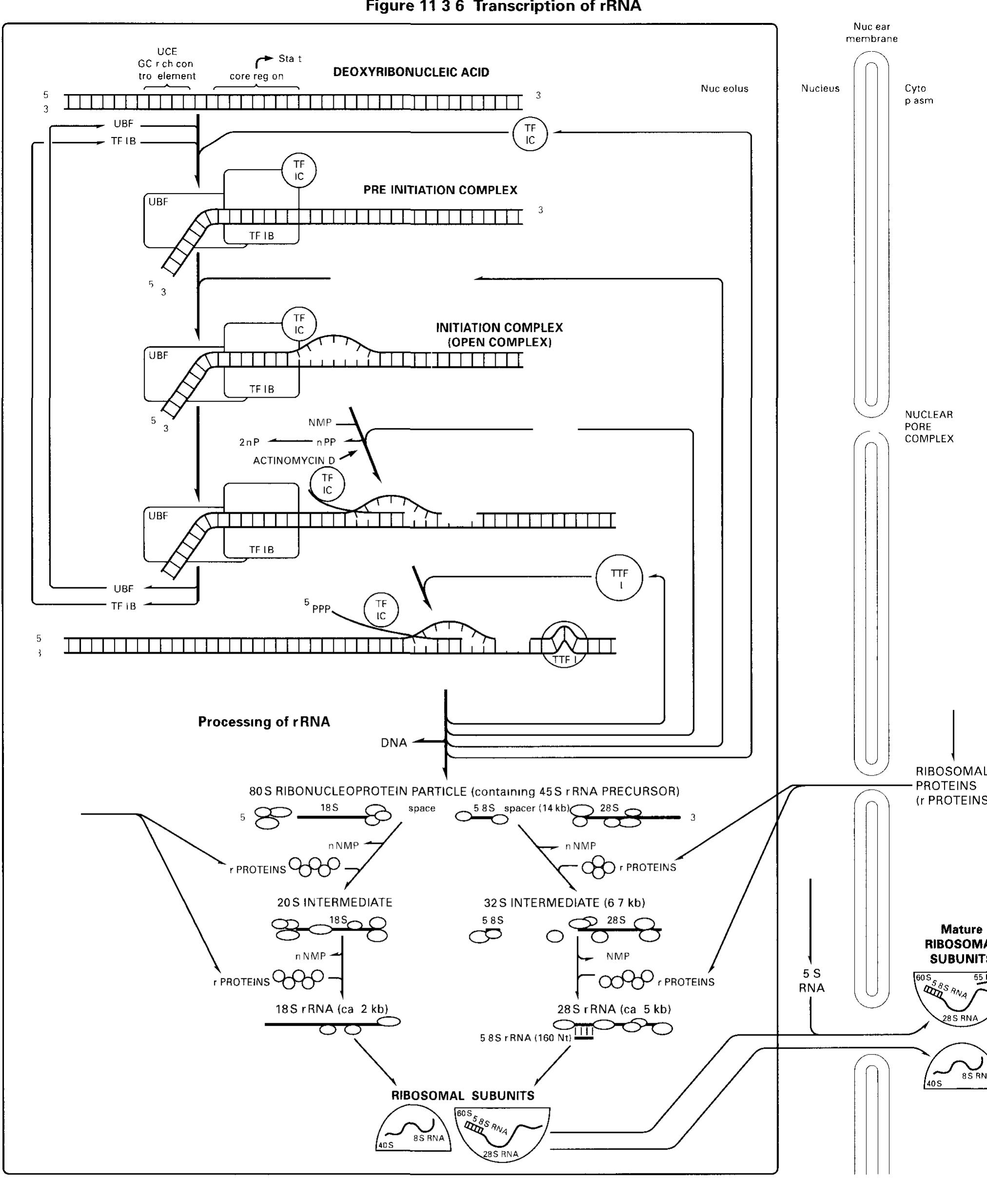






![RNA polymerase I core promoters (Fig. 11.4-3): RNA poly- merase I promoters (and the vast majority of RNA polymerase III] promoters) lack TATA boxes The core promoter (in mammals and amphibians) consists of two critically spaced sequences the upstream control element (UCE, location at -200 -—100 nucleo tides from the start site) and the core region (location at-50 +20 nucleotides therefore enclosing the start site) Only little tran scriptional control 1s exerted on pre-rRNA synthesis](https://figures.academia-assets.com/39117587/figure_190.jpg)

















![Infection of the bacterial cell (Figure 12.2-3): Adsorption to the E colt host cell takes place via specific interaction of the phage tai] fiber and a maltose group of the bacterial outer membrane Then the phage DNA 1s injected through the tail into the host cell The linear DNA 1s transformed into a cyclic form, whereupon the host DNA ligase closes the phage genome ring covalently and the DNA gyrase produces supercoiled phage DNA Now the phage enters one of two different states](https://figures.academia-assets.com/39117587/figure_203.jpg)



![The initiation complex for reverse transcription [consisting of the viral enzyme reverse transcriptase (RT), the nucleocapsid pro- teins and the viral primer t-RNA] 1s being activated by the viral](https://figures.academia-assets.com/39117587/figure_207.jpg)






![[pb glucuronate (2 sulfate) (a1— 4) N sulfo b glucosamine (6 sulfate) (&@1— 4)] (mostly) HEPARIN 6* 10> 25*10'Da contains also some D-Gal and b xylose Large quantities are found in mast cells lung and liver Heparin ts an in hibitor of blood coagulation (20 3) by promoting complex forma tion between active proteases (Ila [Xa, Xa) and antithrombin II]](https://figures.academia-assets.com/39117587/figure_213.jpg)








































![Figure 15.6-2 Redox Potentials E4[mV, 25 °C, pH = 7 0] of Substrate Couples](https://figures.academia-assets.com/39117587/figure_240.jpg)










































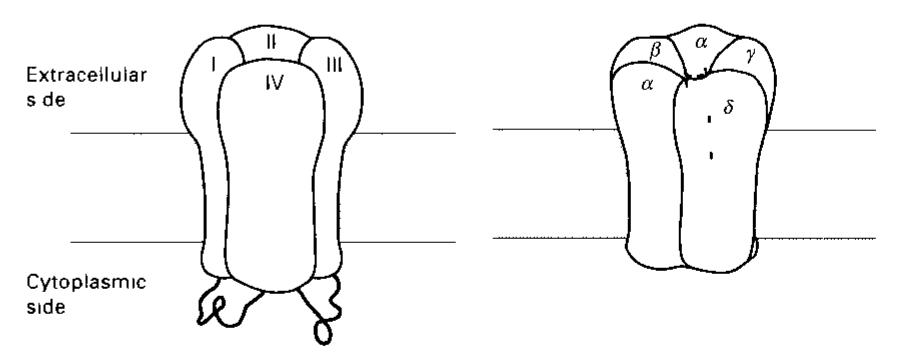







![Table 17.4-2. Mammalian Gg, Proteins (Tightly Associated Complexes) Membrane anchored_ * by farnesy] at C terminal serine # by geranylgeranyl at C terminal leucine Also known B, B; ¥,(# pairs with B, and 8B, widespread occurrence) y;(# pairs with B and B widespread occurrence) y; (# pairs with B, and B) widespread occurrence) ys # in olfactory epithelium) y , G# pairs with B, and B.) y (* similar to y, pairs with B widespread occurrence)](https://figures.academia-assets.com/39117587/table_085.jpg)